本网讯(通讯员 王超楠 图片 王超楠)2025年5月5日,我院李洪连教授团队/张超教授课题组在植物学权威期刊《Plant Biotechnology Journal》(IF2024=11.2)在线发表了题为“A fijivirus capsid protein hijacks autophagy degrading an ω-3 fatty acid desaturase to suppress jasmonate-mediated antiviral defence”的研究论文。该研究通过代谢组学分析发现,水稻黑条矮缩病毒(rice black-streaked dwarf virus,RBSDV)侵染后,水稻叶片中不饱和脂肪酸(Unsaturated fatty acid, UFAs)含量显著降低,其中茉莉酸(jasmonic acid, JA)生物合成前体α-亚麻酸(C18:3)的减少尤为突出。进一步研究表明,RBSDV外壳蛋白P10能够直接与水稻脂肪酸去饱和酶7(fatty acid desaturase 7, FAD7)相互作用,并通过自噬途径促进OsFAD7的降解,从而抑制C18:3及JA的积累,削弱植物的抗病防御反应。本研究揭示了病毒通过劫持自噬系统干扰脂肪酸代谢,进而抑制JA信号通路的分子机制,不仅深化了对病毒-寄主互作的理解,也为抗病毒作物育种和新型抗病毒策略的开发提供了重要的理论依据。

细胞自噬作为一种高度保守的蛋白质降解机制,在植物-病毒互作过程中发挥关键作用。已有研究表明,自噬在植物抗病毒防御中具有双重功能:一方面,寄主可通过自噬途径选择性降解病毒蛋白或病毒颗粒,从而抑制病毒侵染;另一方面,某些病毒已进化出劫持寄主自噬系统的策略,通过干扰寄主防御反应或促进病毒颗粒组装来增强其致病性。植物细胞中,自噬降解途径通常依赖于自噬相关蛋白8(autophagy-related proteins 8, ATG8)与底物蛋白中的ATG8相互作用基序(ATG8-interaction motif, AIM)(保守序列W/F/Y-X-X-L/I/V, X表示任意氨基酸)的特异性识别。本研究通过生物信息学分析,发现在OsFAD7中存在两个潜在的AIM基序。蛋白构象模拟显示,AIM1位点的突变对OsFAD7的空间构象产生显著的影响(图1a)。进一步的分子互作实验证实,当AIM1突变后,OsFAD7与多个OsATG8的相互作用明显受到影响(图1b-c),且GFP-OsFAD7-mAIM1突变体能够抵抗OsATG8介导的蛋白降解,而AIM2突变则不影响这一过程(图1d)。这表明AIM1基序是OsATG8识别和降解OsFAD7的关键位点。此外,AIM1基序同样对P10促进OsFAD7的降解至关重要(图1e),而且P10能够显著增强OsATG8介导的OsFAD7的降解(图1f)。这一系列发现充分证明,RBSDV外壳蛋白P10通过AIM1-OsATG8这一特异性识别系统来促进OsFAD7的降解,揭示了病毒蛋白劫持寄主自噬途径促进自身侵染的分子机制。
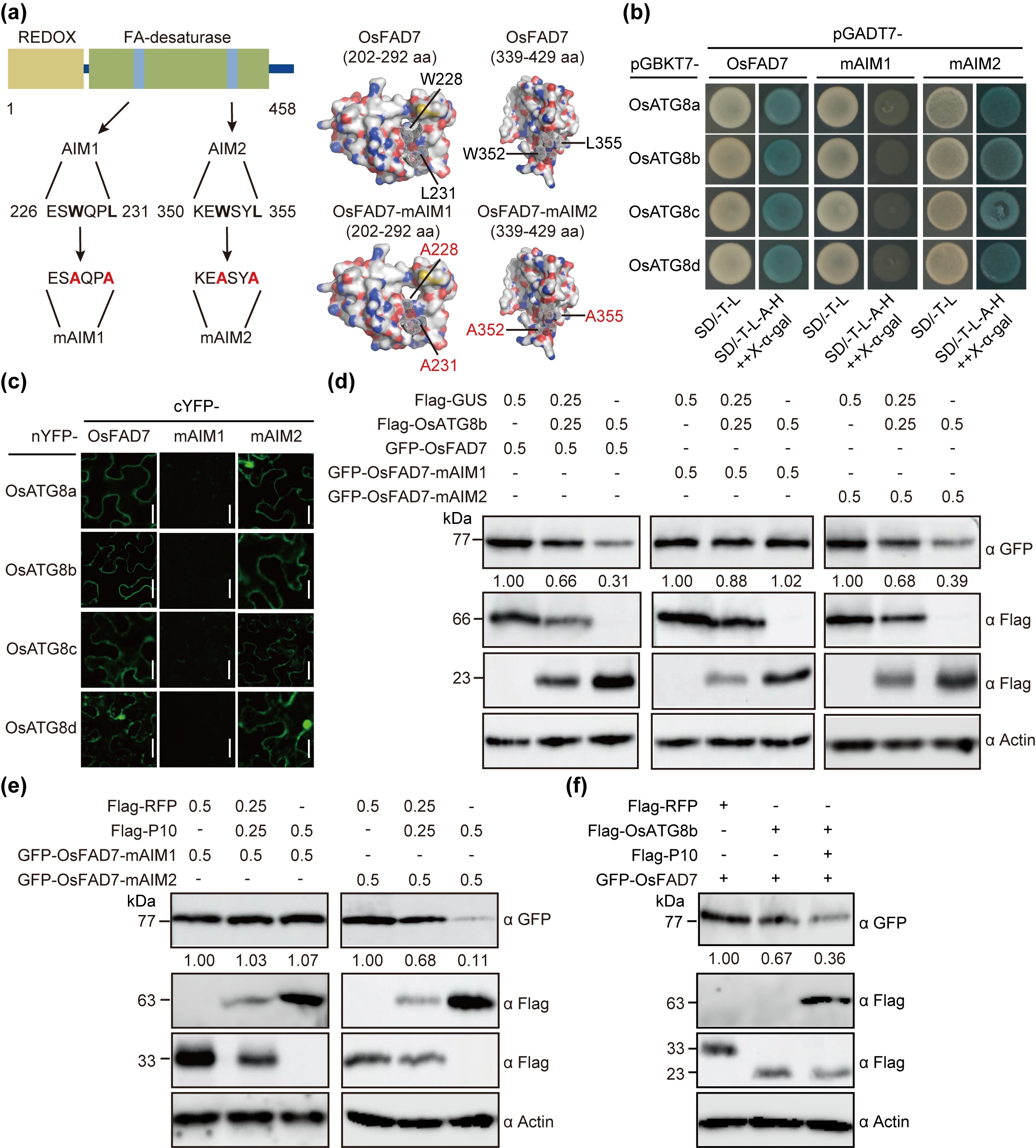
图1 RBSDV P10促进OsATG8介导的OsFAD7的自噬降解
河南农业大学与长江大学联合培养博士研究生刘俭俭和河南省农科院敬新欣博士为论文的共同第一作者,我院张超教授、青年教师王超楠博士和长江大学章松柏教授为该研究工作的共同通讯作者。李洪连教授、青年教师李彭拜博士以及福建农林大学吴祖建研究员对本研究提供重要支持。研究工作得到了河南省自然科学基金(252300420676)、中国博士后面上项目(2023M731005)、河南农业大学高层次人才专项支持基金(30500948)和校企合作开发基金(30803130)等项目的资助。
论文链接:https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.70119




